タンパク質のリン酸化を抑制することで損傷した視神経の再生を促進
外力による損傷以外の緑内障などの疾患への有効性検証に期待
発表のポイント
- 脳や脊髄などの中枢神経系は、一度ダメージを受けると再生しにくく、治療が難しい。
- 研究グループは、神経の軸索の形態を支えるタンパク質の一つであるCRMP2のリン酸化を抑制することが中枢神経の一部である視神経の再生に有効であることを明らかにした。
- 患者数の多い緑内障などでも外力による視神経損傷と同様の病態が起きている可能性があり、今後の検証が期待される。
早稲田大学理工学術院の大島登志男(おおしまとしお)教授、国立研究開発法人国立精神・神経医療研究センター神経研究所の荒木敏之(あらきとしゆき)部長、公立大学法人横浜市立大学大学院医学研究科の五嶋良郎(ごしまよしお)教授らの研究グループは、外力によって損傷した視神経の再生が、CRMP2 (collapsin response mediator protein)というタンパク質のリン酸化を抑制することで促進されることを明らかにしました。
中枢神経系は、一度ダメージを受けると再生しにくく、これが神経疾患の治療を難しくしています。本研究グループは、中枢神経系が損傷を受けた際に神経細胞側でリン酸化するCRMP2というタンパク質に着目し、これまでも脊髄の損傷モデルで、独自に開発したCRMP2のリン酸化を抑制した遺伝子改変マウスは、野生型のマウスよりも運動機能回復が良好であることを報告してきました。今回の研究では、視神経の損傷モデルでこの遺伝子改変マウスを用いて、CRMP2のリン酸化抑制が損傷後の神経軸索再生に有効かを検討し、神経再生が顕著に促進されていることを見出しました。
今後、緑内障などの疾患に対しても、CRMP2のリン酸化抑制が病気の進行を抑制できるか遺伝子改変マウスを用いて検証するとともに、CRMP2のリン酸化を抑制する薬剤の同疾患への有効性を検証することが期待されます。
なお、本研究成果は、英国のオンライン科学雑誌『Scientific Reports』に2019年5月10日午前10時(現地時間)に掲載されました。
これまでの研究で分かっていたこと(科学史的・歴史的な背景など)
脳や脊髄などの中枢神経系は、一度ダメージを受けると再生が起こらず、これが神経疾患の治療を難しくしている。再生には、神経細胞が新たに作られる神経新生と、一度切断あるいは損傷を受けた神経の軸索が再び伸長する神経軸索再生がある。大人の脳での神経新生(成体神経新生)はヒトなど哺乳類では脳の限られた部位でのみ起きていて、脳の一部の障害の回復に寄与している可能性が報告されているが、その増殖や分化の制御のメカニズムは不明な点が多い。成体神経新生の研究が盛んに行なわれているのは、その増殖や神経細胞への分化を制御するメカニズムを利用して、神経再生を促すことが出来る可能性があるからである。一方の神経軸索再生が中枢神経系で起こりづらいのは、それを阻害する仕組みがあるからである。脊髄損傷では加えて損傷部の瘢痕形成などの物理的障害が問題となる。中枢神経系の一部である視神経も損傷を受けると、瘢痕形成は顕著でないものの、再生が難しいが、その原因には再生を阻害する因子の存在とその阻害因子に対する神経細胞側の反応がある。こうした阻害因子やその神経細胞側の反応のメカニズムが次第に明らなになりつつあり、それを利用した治療法の開発が試みられている。
今回の研究で新たに実現しようとしたこと、明らかになったこと
神経細胞側の阻害因子に反応して軸索の伸長が抑制される仕組みの中で、軸索の形態を支える骨格タンパク質の微小管の不安定化がある。微小管はa-チュ―ブリンとb-チューブリンが2量体を形成して、それが重合して連なった構造をしており、重合と脱重合により微小管の伸長と退縮が起こる。こうした構造を安定化するものに、微小管結合タンパク質がある。今回の研究で着目しているCRMP2 (collapsin response mediator protein)は、微小管結合タンパク質の一つで、非リン酸化状態で微小管に結合して微小管を安定化するとともに、その重合を促進することが知られている。本研究グループはこれまでにCRMP2がリン酸化を受けると微小管との結合能が失われ、結果として微小管が不安定化して軸索の退縮が起きることを報告している(Uchida et al., 2005)。このCRMP2のリン酸化による軸索の退縮は軸索伸長を阻害する因子であるSema3A*1で起きるが、CRMP2はこの反応を伝えることから命名されている(collapsinはニワトリのSema3Aの旧名)。中枢神経系が損傷を受けた際には、このSema3Aに加えて、軸索を取り巻くミエリン由来のミエリン関連阻害因子と呼ばれる一連の阻害因子が軸索再生を抑制するが、軸索側ではCRMP2のリン酸化が起きており、そのリン酸化を抑制することで神経再生を促そうというのが今回の研究の目的である。こうした目的のため、本研究グループはこれまでにCRMP2のリン酸化を抑制した遺伝子改変マウスを開発している(Yamashita et al., 2012)。CRMP2は何種類かのキナーゼによりリン酸化されるが、本研究グループが着目しているのはSer522のサイクリン依存性キナーゼ5(Cdk5)によるリン酸化で、このリン酸化依存的にさらに別のキナーゼであるGSK3bによりThr509, Thr514, Ser518の3つの部位がリン酸化を受けるが(Uchida et al., 2005)、この522番目のSerをAlaに置換してリン酸化されなくしたのが本研究グループの開発したCRMP2KI (ノックイン (knock-in, KI))マウスである(Yamashita et al., 2012)。このマウスでは、4箇所のリン酸化が阻害されることとなり、Sema3Aなどの阻害因子による軸索再生阻害を受けないことが予想される。本研究グループはこれまでに、脊髄損傷のモデル系で、CRMP2KIマウスが野生型に比べて損傷からの運動機能回復が良好であることを報告している(Nagai et al., 2016)。今回の研究では、視神経の損傷モデルでCRMP2のリン酸化抑制が損傷後の神経軸索再生に有効である可能性を検討している。今回の研究で、以下の点が明らかになっている。
- CRMP2KIマウスでは視神経損傷直後に起きる微小管の脱重合が軽度であること。
- CRMP2KIマウスでは網膜にある視神経の元になる細胞(網膜神経節細胞)の脱落が少ないこと(図1)。
- 視神経損傷4週間後に比較するとCRMP2KIでは神経再生マーカーのGAP43の発現が増加していること(図2)。
- 視神経再生をトレーサーを用いて調べると*2、CRMP2KIでは神経再生が顕著に促進されていることが明らかとなった(図3)。
そのために新しく開発した手法
前述の通り、遺伝子改変によりCRMP2KIマウスを開発している(Yamashita et al., 2012)。開発は、今回の共同研究者である横浜市立大学大学院医学研究科の五嶋良郎教授との共同研究で行なった。
研究の波及効果や社会的影響
視神経損傷は、物理的外力によるダメージで起きるものであるが、より患者数の多い眼科疾患である緑内障などの病態でも同様のメカニズムで神経再生阻害が起きている可能がある。本研究に加え、本研究グループの脊髄損傷モデルにおけるCRMP2のリン酸化を抑制することが、中枢神経系の再生を促進するという結果(Nagai et al., 2016)や、CRMP2KIではALSモデルマウスの症状の進行が抑えられる(Numata-Uematsu et al., 2019)といった研究成果は、CRMP2のリン酸化抑制をターゲットとした薬剤が中枢神経系の神経再生促進に有用である可能性を強く示唆するものである。実際、本研究グループはCRMP2のリン酸化を抑制し、CRMP2と結合するタンパク質との結合を変化させる薬剤であるLKEが脊髄損傷モデルの機能回復に有用であることを報告している(Kotaka et al., 2016)。
今後の課題
今回の研究成果は、遺伝子改変したマウスでの結果であり、前述のLKEなどCRMP2のリン酸化を抑制する薬剤が神経再生に有用であることを検証する必要がある。また、より患者数の多い緑内障などでも外力による視神経損傷と同様の病態が起きている可能性があり、緑内障などの眼科疾患にたいしてもCRMP2のリン酸化抑制が病気の進行を抑制できるか、CRMP2KIマウスを用いて検証するとともに、CRMP2のリン酸化を抑制する薬剤の同疾患への有効性を検証することが期待される。ALSなどの神経系変性疾患に対してもCRMP2のリン酸化抑制が有効であるか検証する必要がある。最近本研究グループは、MPTPという薬剤でドーパミン神経細胞死を誘導したパーキンソン病モデルマウスにおいて、軸索変性がCRMP2のリン酸化抑制により軽減されることを報告している(Togashi et al., 2019)。従って、CRMP2のリン酸化抑制が神経損傷及び神経変性疾患に対する治療ターゲットとなることが期待され、今後の検証が期待される。
用語解説
- *1 Sema3A
Semaphorin 3Aの略で、神経軸索の退縮を引き起こす典型的な分子として知られている。神経細胞のニューロピリンとプレキシンという受容体を介してその反応が細胞で引き起こされる。発見当初は種により様々な名称、例えばニワトリではcollapsinと呼ばれていた。この反応を伝える細胞内のシグナルタンパク質としてcollapsin response mediator protein (CRMP)が同定されている(Goshima et al., 1996)。
- *2 トレーサーによる標識
硝子体中にトレーサーを注入することにより、トレーサーが網膜の神経節細胞に取り込まれ、順行性輸送によりその軸索である視神経の末端まで運搬されるため、軸索を含めた神経細胞全体を標識することができる。
論文情報
- 雑誌名:Scientific Reports
- 論文名:Genetic inhibition of CRMP2 phosphorylation at serine 522 promotes axonal regeneration after optic nerve injury
- 執筆者名:Shunsuke Kondo1, Kazuya Takahashi1, Yuki Kinoshita1, Jun Nagai1, Shuji Wakatsuki2, Toshiyuki Araki2, Yoshio Goshima3, Toshio Ohshima1
※1所属:早稲田大学先進理工学研究科生命医科学専攻
※2所属:国立研究開発法人国立精神・神経医療研究センター 疾病研究第5部
※3所属:公立大学法人横浜市立大学 大学院医学研究科 分子薬理神経生物学
- 掲載日時:2019年5月10日午前10時(イギリス現地時間)
- 掲載URL:https://www.nature.com/articles/s41598-019-43658-w
- DOI:10.1038/s41598-019-43658-w
研究助成
- 科研費基盤研究C「CRMPをターゲットとした神経損傷・神経変性疾患治療戦略開発」(研究代表者:大島登志男)
- その他、JST・CREST、文部科学省「先端融合領域イノベーション創出拠点形成プログラム」
参考図
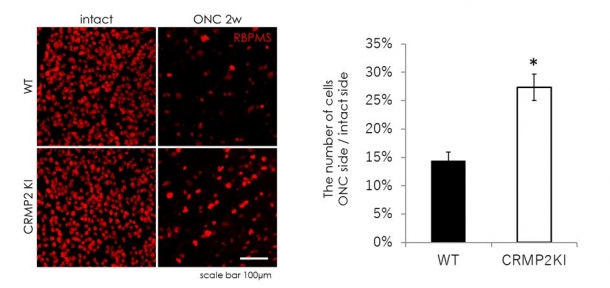
図1 野生型(WT)とCRMP2KIマウスの視神経損傷2週間後(ONC 2W)の網膜を比較すると、CRMP2KIマウスで赤く染色された神経節細胞の数が多いことが分かる。グラフは、非損傷側(intact side)に比べた損傷側(ONC side)の細胞のパーセントを示している。
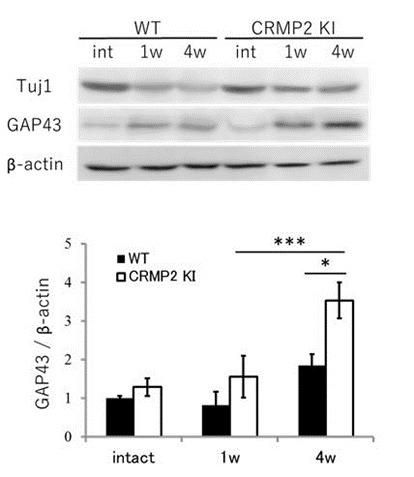
図2 野生型(WT)とCRMP2KIマウスの視神経の損傷前(int)、損傷1週間後(1W)、4週間後(4W)の各種タンパク質の量を比較したもの。再生した神経のマーカーGAP43がCRMP2KIマウスの損傷後の視神経で多いことが分かる。
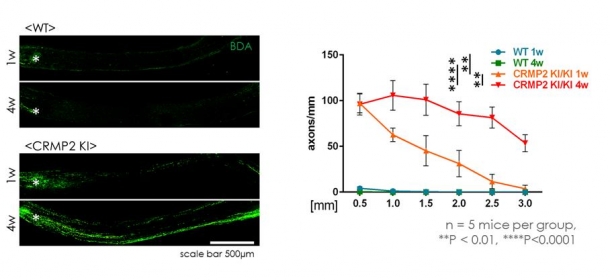
図3 トレーサーによる標識の結果、CRMP2KIの損傷4週間後(4W)で標識された軸索が多く観察される。損傷部位からの一点距離で標識された軸索の数を数値化したグラフからも、CRMP2KIでは損傷後の視神経再生が顕著に起きていることが分かる。






