地球規模の急激な寒冷化が酵素の進化を促進
40億年に渡る生物と地球環境の共進化の謎に迫る
発表のポイント
- 共通祖先酵素から、現存の常温菌酵素をつなぐ進化経路上の11個の中間祖先酵素を復元したところ、高温に適した触媒特性から低温に適した特性への大きな変化は、21~25億年前の2つの連続した中間祖先の間で起こることを発見しました。
- 低温に適した酵素では、高温に適した酵素では見られない活性部位が閉じた構造が存在し、開閉遷移のエネルギーコストと触媒反応に必要な活性化エネルギーが低減されることで、低温での触媒反応がより効率的になりました。
- 今回の研究手法は、さまざまな酵素に適用することができ、生物とその酵素がどのように40億年に渡る地球環境の変化に適応し進化してきたかを明らかにすることができると考えられます。これは持続可能な社会の実現に向けた技術革新へと繋がると期待されます。
早稲田大学人間科学学術院の赤沼 哲史(あかぬま さとし)教授、八木 創太(やぎ そうた)講師、同大学院人間科学研究科修士課程(当時)の崔 爽(さい そう)および理化学研究所の田上 俊輔(たがみ しゅんすけ)チームリーダー、Subrata Dasgupta研究員らの研究グループは、好熱性の祖先酵素から常温性である大腸菌酵素への進化を解明するため、11段階の中間祖先酵素を再現し、活性の変化を解析しました。その結果、約21~25億年前に低温で活発に働く性質が大きく向上したことが分かりました。当時の地球は温室効果ガスの減少による氷河期にあったことから、急激な寒冷化が酵素の低温適応進化を促した可能性があります。今回の解析結果は、気候変動が生命の進化にどのような影響を与えてきたかを示す重要な証拠であり、地球史と生物進化の関係を理解する上で新たな知見をもたらしました。酵素の温度適応メカニズムを詳細に理解することで、エネルギー効率の高い触媒反応や、環境負荷を低減した工業プロセスの開発にも繋がることが期待されます。
本研究成果は『Protein Science』(論文名:Insights into the low-temperature adaptation of an enzyme as studied through ancestral sequence reconstruction)にて、2025年2月19日(水)にオンラインで掲載されました。
(1)これまでの研究で分かっていたこと
何十億年もの間、酵素はその酵素を持つ生物の生息環境の変化に応じて進化してきました。たとえば、高温環境に生息する好熱性生物※1の酵素は、熱に強く高温下での反応が非常に活発ですが、低温ではその働きが大きく低下することがわかっています。一方、一般的な中温性生物※1や好冷性生物※1から得られる酵素は、熱に対する耐性は不十分でありながら、低温で高い触媒活性を示す傾向があります。様々な証拠から、最古の原始生物は高温環境に生息し、高温環境に適応した酵素を持っていたことが示唆されています。したがって、現存する中温性の酵素や好冷性酵素は、地球の表面温度の低下に伴い、進化の過程で低温に適応してきたと考えられています。中温性、好冷性の酵素が低温で活発に働くのは、酵素表面の柔軟性と密接に関連していると考えられていますが、具体的なメカニズムやアミノ酸配列との関係はまだ解明されていません。
以前の研究では、現存する相同アミノ酸配列の解析をもとに、真正細菌※2の共通祖先に由来する2種類の3-イソプロピルリンゴ酸脱水素酵素(IPMDH)※3を、祖先配列再構成(Ancestral Sequence Reconstruction, ASR)技術※4を用いて再現しました。ASRとは、分子系統解析※5と遺伝子工学・タンパク質工学の技術を組み合わせることによって、過去に存在したが現在は消失してしまったタンパク質(酵素)の祖先アミノ酸配列を推定し、再現する技術です。ASRによって再現された祖先IPMDHは、非常に高い熱安定性を示しましたが、中温性である大腸菌が持つIPMDH(EcIPMDH)と比較すると、低温環境下での触媒活性が著しく劣ることが明らかになっています。
(2)今回の研究で新たに実現しようとしたこと、明らかになったこと
今回の研究では、好熱性である最古の共通祖先※6の酵素から、中温性である大腸菌が持つ酵素へと至る進化の過程で、どのように低温活性を改善してきたのかを探ることを目的としました。具体的には、真正細菌の共通祖先IPMDHとEcIPMDHをつなぐ進化の軌跡に沿って、その間の11個の中間祖先酵素を再現し(図1)、各段階での酵素活性の変化、特に低温活性の変化を詳しく解析しました。
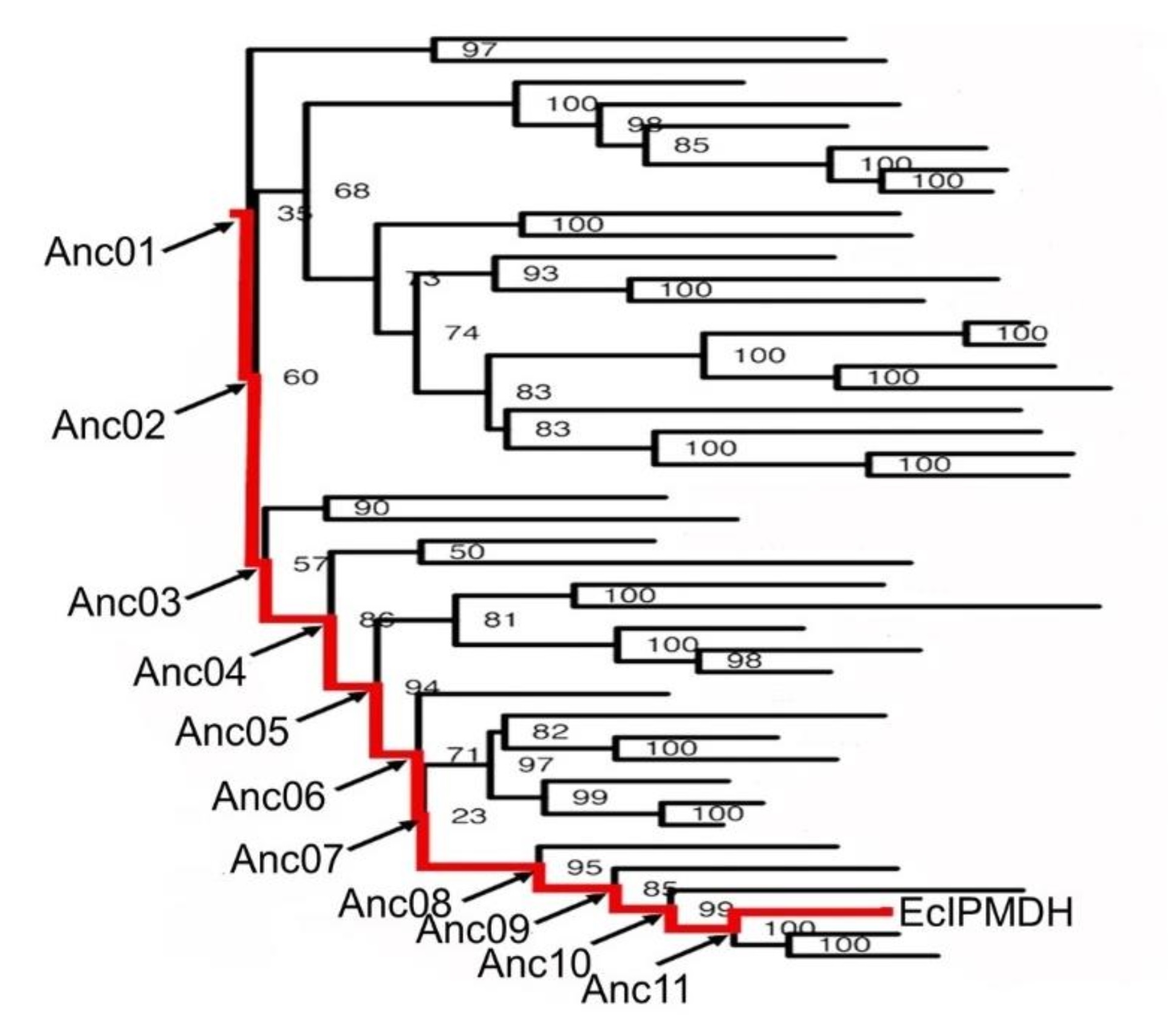
図1:真正細菌が持つIPMDHのアミノ酸配列から構築された分子系統樹。真正細菌共通祖先IPMDHと大腸菌IPMDH(EcIPMDH)をつなぐ進化経路を赤線で示し、その経路上の11個の中間体の位置を表示した。系統樹内の数値はブートストラップ確率を表す。
25℃における触媒活性の大きさは、最古の祖先からEcIPMDHへの進化の過程で直線的な変化を示さず、5番目の中間祖先(Anc05)と6番目の中間祖先(Anc06)の間で25℃における触媒活性の劇的な向上が観察されました(図2)。
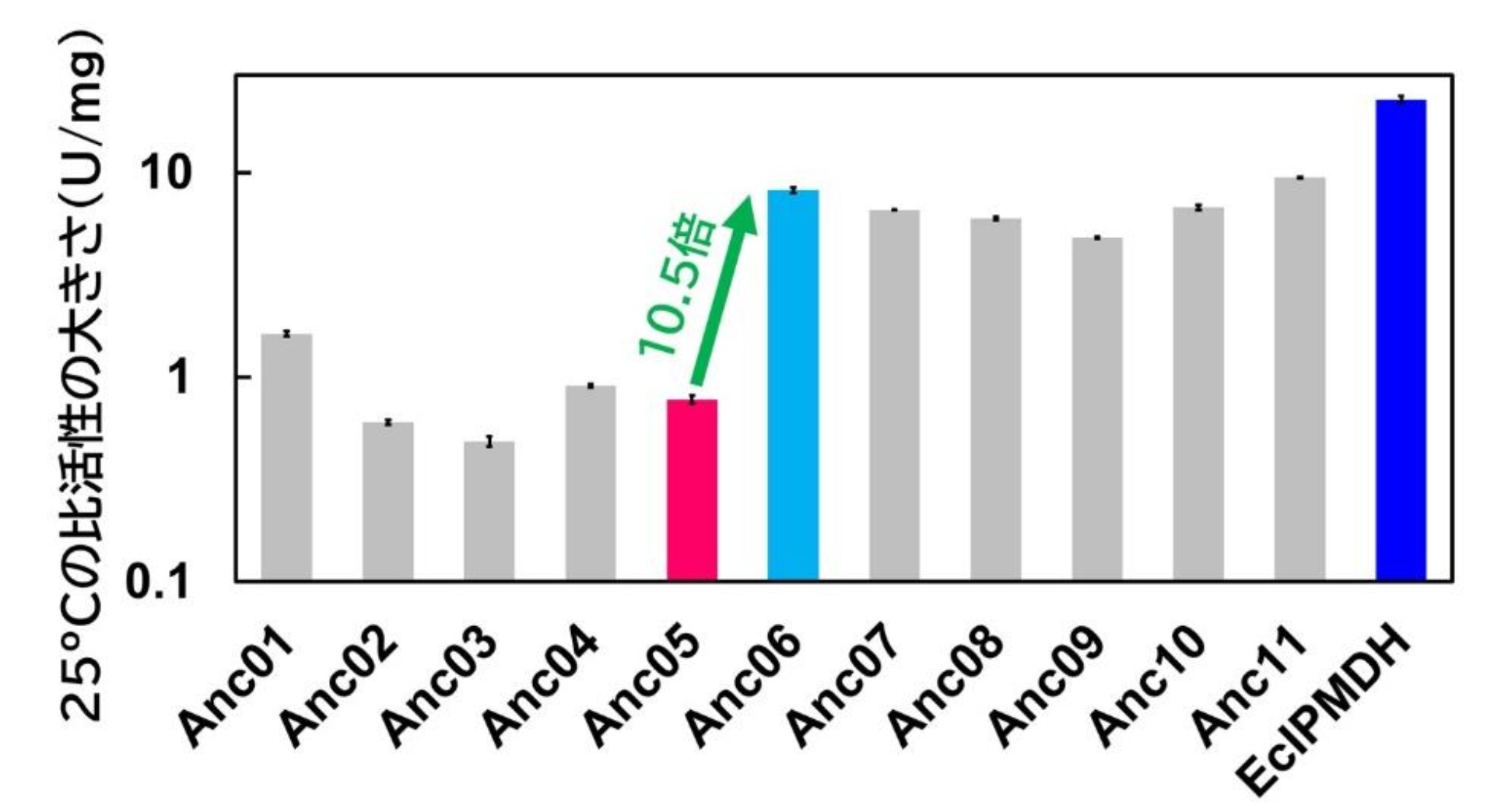
図2:25℃における各中間祖先IPMDHと大腸菌IPMDH(EcIPMDH)の比活性の大きさ。縦軸は対数目盛を使用。各値は3回の測定値の平均であり、エラーバーは標準誤差を表す。Anc06の活性はAnc05と比較して10.5倍向上した。
さらに、中間祖先酵素間の配列比較と部位特異的突然変異解析の組み合わせにより、低温触媒活性を増強する3つの重要なアミノ酸置換を同定しました。これまでは、酵素の温度適応は主に活性部位残基の突然変異によって引き起こされると考えられてきました。しかし、本研究で同定された3つの重要なアミノ酸置換は、いずれも活性部位から離れた位置で起こっていました(図3)。この発見は、ASRが酵素の活性部位から離れた場所に位置する、低温活性の向上に重要なアミノ酸残基を同定する方法であることも示しました。さらに興味深いことに、同定した3つのアミノ酸置換のうち、低温での触媒活性に最も大きな影響を与えるアミノ酸置換は、酵素の耐熱性には顕著な影響を与えませんでした。しかし、この置換は触媒反応の活性化エネルギー※7を著しく低下させることで、低温活性を大きく向上させることが分かりました。
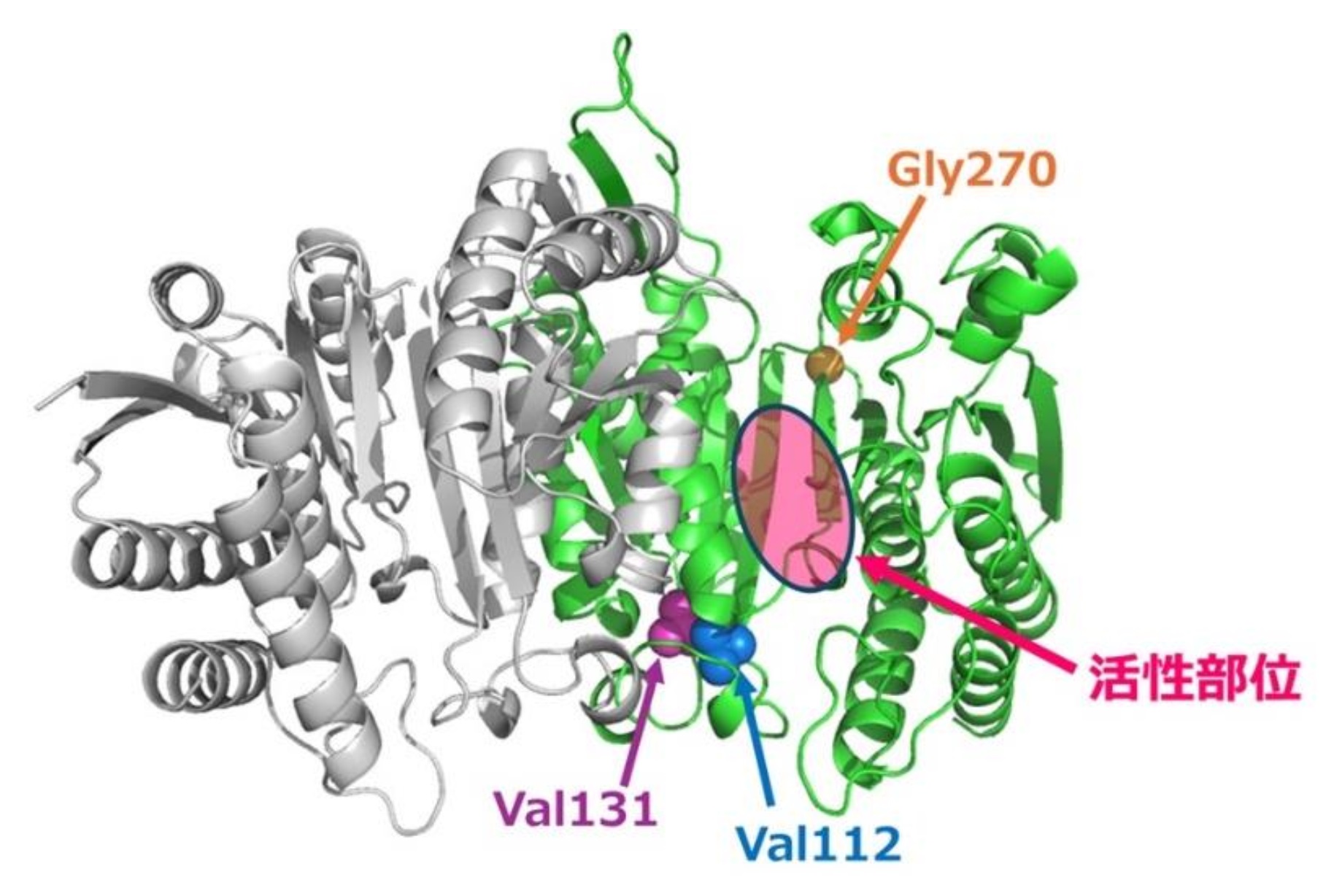
図3:Alphafold2を用いて予測したAnc05の二量体構造のモデル。2つのサブユニットを異なる色で示した。Anc05のVal112をPhe, Val131をPhe, Gly270をAlaに置換すると25℃の比活性が大きく向上する。これら3つの低温高活性化に重要なアミノ酸置換部位は活性部位から離れた場所で生じた。Val112とGly270は、2つのドメインをつなぐβストランドの端に位置し、Val131の側鎖はVal112の側鎖と相互作用している。
さらに、Anc05とAnc06の間で観察された触媒特性の明確な違いから、その進化の間に地球規模の大きな環境変化があったか探りました。祖先配列の再現に使用した分子系統樹に、生物種間の分岐年代の情報を組み込んで、それぞれの中間祖先酵素のおおよその年代を推定しました(表1)。その結果、29億年以上前に存在したと推定されたAnc01からAnc03で観察された高い耐熱性は、元素同位体分析※8のデータから予測された、当時の温暖な地球気温と一致しました。対照的に、その後の中温性への変化は、長期的な地球環境の冷却傾向を反映していると言えます。特に、好熱性のAnc05から中温性のAnc06への進化は、25億年前から21億年前の間に起こったと推定されましたが、この時期には、大酸化現象に伴う温室効果ガスである大気中のメタンガスの劇的な減少によって、地球規模の氷河期があったと考えられています。さらに、「スノーボール・アース」※9との関連も指摘されています。つまり、全球規模の急激な寒冷化が、Anc06で観察された酵素の低温適応進化をもたらしたと考えられます。
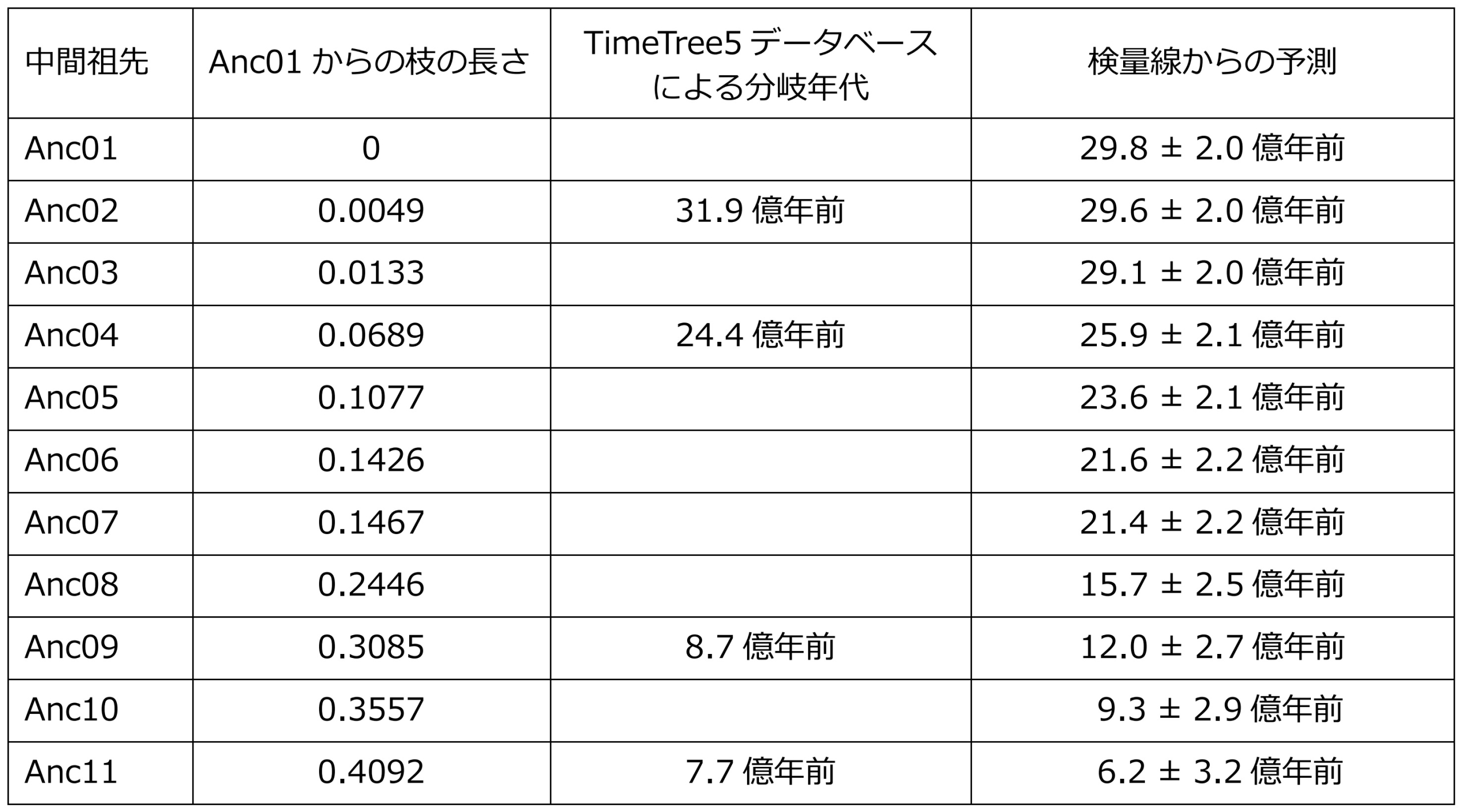
表1:中間祖先IPMDHが存在した年代予測。Anc02、Anc04、Anc09およびAnc11の分岐年代はTimeTree5データベース(http://www.timetree.org/)から得た。他の中間祖先の年代は系統樹の枝の長さと検量線を用いて概算した。
IPMDHは触媒過程で活性部位が開いた構造から、基質や補酵素の結合により閉じた構造へと遷移することが知られています。分子動力学シミュレーション※10により、Anc05とAnc06の構造分布を比較しました。初期構造からの構造のずれを示すα炭素位置の二乗平均平方根偏差(CαRMSD)と慣性半径(Rg)のヒートマップを比較したところ(図4)、Anc05は主に開いた形の周辺で揺らぎを示したのに対し、Anc06はそれぞれ開いた形と部分的に閉じた形に対応する2つの頻度ピークが観察されました。低温で活発に働くAnc06に閉じた形が存在することで、開型から閉型への遷移のエネルギーコストと、触媒反応に必要な全体的な活性化エネルギーが低減され、低温での触媒反応がより効率的になったと考えられます。
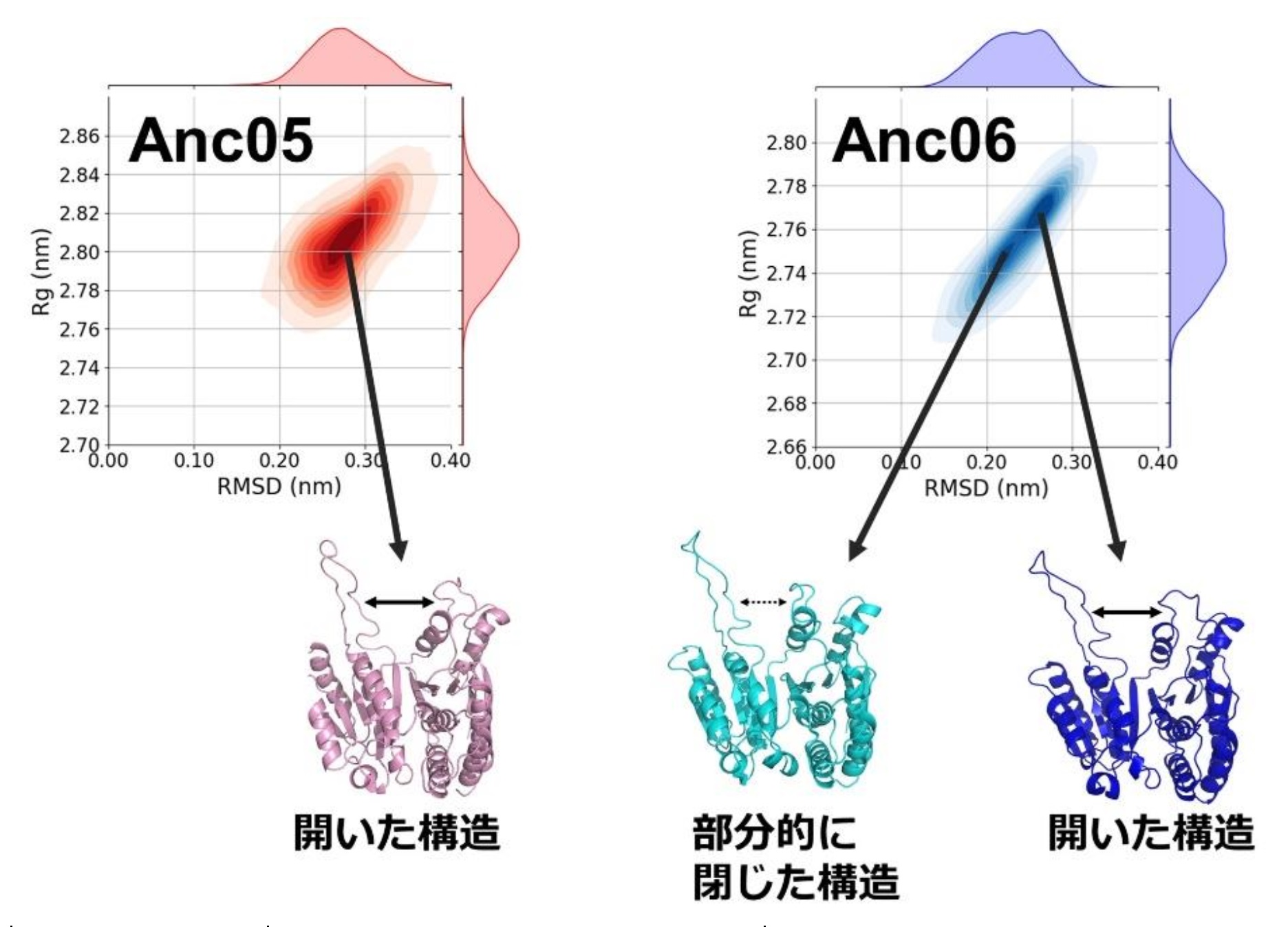
図4:分子動力学シミュレーションにより観察されたAnc05とAnc06の立体構造アンサンブルの初期構造からのずれ(RMSD、x軸)と慣性半径(Rg、y軸)の2次元度数分布。ヒートマップはサンプリングされた構造の密度を表し、色が濃いほど頻度が高いことを示している。RMSDとRg値の周辺度数分布を、それぞれヒートマップの上と右に表示している。頻度のピークに対応する代表的な構造スナップショットをヒートマップの下に示した。Anc05の主な構造は開状態に対応するのに対し、Anc06では開いた構造と部分的に閉じた構造の両方が観察される。
(3)研究の波及効果や社会的影響
本研究では、好熱性の共通祖先から現存する常温菌酵素へと至る進化過程を再現し、低温適応のメカニズムを分子レベルで解明しました。その結果、約21~25億年前の氷河期において、地球規模の急激な寒冷化が酵素の低温活性化を促したことが明らかになりました。この発見は、気候変動が生命の進化にどのような影響を与えてきたかを示す重要な証拠であり、地球史と生物進化の関係を理解する上で新たな知見をもたらしました。さらに、酵素の温度適応メカニズムを詳細に理解することで、低温でも高い活性を維持できる酵素の設計が可能となり、エネルギー効率の高い触媒反応や、環境負荷を低減した工業プロセスの開発につながることが期待されます。
本研究成果は、進化生物学、地球化学、タンパク質工学といった異分野の連携を促進し、学際的研究の発展に貢献することが期待できます。特に、生命の進化に関する理解を深めるとともに、持続可能な社会の実現に向けた技術革新へとつながることが期待されます。
(4)今後の課題
今後は低温適応の分子メカニズムをより詳細に解明する必要があります。また、温度環境に限らず、酸性・アルカリ性、高圧など、異なる環境因子が酵素の進化にどのような影響を与えるのかを明らかにし、環境適応の一般的な原理を探ることも求められます。さらに、生命の進化における環境適応の全体像の理解には、1種類の酵素だけでなく代謝全体に着目することも必要となります。加えて、過去の気候変動と生物進化の関係を明らかにできれば、現代の気候変動が生態系や進化に与える影響を予測する上でも有益な知見となるはずです。こうした進化メカニズムの理解は、産業分野にも波及効果があり、低温でも高い活性を持つ酵素を設計することで、エネルギー消費を抑えた触媒反応や環境負荷の低減など持続可能な社会の構築への貢献が期待されます。
(5)研究者のコメント
本研究では、好熱性祖先から常温菌酵素への進化を再現し、地球規模の急激な寒冷化が酵素の低温高活性化を促したことと、その分子メカニズムを明らかにしました。今後は過去の気候変動と進化の関係を明らかにするだけでなく、将来の生態系への影響予測にも取り組めたらと思います。また、低温適応型酵素の設計により、省エネルギー型産業プロセスや持続可能な社会の実現にも貢献したいと考えています。
(6)用語解説
※1 好熱性生物、中温性生物、好冷性生物
好熱性生物とは主に50℃以上の高温で生育する生物であり好熱菌とも呼ぶ。その中でも、生育上限温度が70℃を超える微生物を高度好熱菌、最適な生育温度が80℃以上の微生物を超好熱菌と呼ぶ。中温性生物とは常温生物とも呼び、最適な生育温度が20℃~50℃である生物を指すことが多い。また、最適な生育温度が20℃未満である生物を好冷性生物と呼ぶ。
※2 真正細菌
地球のすべての生物を、タンパク質合成に関わるRNAの塩基配列の類似性に基づいて分類すると、大腸菌や乳酸菌、藍藻などの微生物が含まれる真正細菌の他に、メタン菌や超好熱性古細菌などが含まれる古細菌と、動物、植物、カビなどが含まれる真核生物の3つのドメインに分けることができる。それぞれのドメインは単系統群となり、それぞれのドメインに属するすべての生物種は、それぞれのドメインにおける共通祖先から進化したと考えられている。
※3 3-イソプロピルリンゴ酸脱水素酵素(IPMDH)
微生物や植物が持つ酵素。アミノ酸の1種であるロイシンの生合成に関わる。ニコチンアミドアデニンジヌクレオチド(NAD)を電子受容体として3-イソプロピルリンゴ酸を酸化的脱炭酸する反応を触媒する。NADから高価な還元型ニコチンアミドアデニンジヌクレオチド(NADH)をリサイクルする過程を含む工業プロセスに利用できる。
※4 祖先配列再構成(Ancestral Sequence Reconstruction, ASR)技術
分子系統解析(※5参照)と実遺伝子工学・タンパク質工学の技術を組み合わせることによって、過去に存在したが現在は既に消失したと想定される遺伝子やタンパク質(酵素)の塩基配列、アミノ酸配列を推定し、再現する方法。現存する生物の遺伝情報やアミノ酸配列を比較し、共通の祖先が持っていたと考えられる配列を計算で推定します。この技術を用いることで、進化の過程を実験的に再現したり、古代生物の性質を調べたりすることができます。
※5 分子系統解析
現存の異なる生物が持つ同一の遺伝子(相同遺伝子)の塩基配列や、異なる生物が持つ同一のタンパク質(相同タンパク質)のアミノ酸配列(相同アミノ酸配列)を比較することによって、それらの遺伝子やタンパク質が、共通祖先遺伝子、あるいは、共通祖先タンパク質からどのような道筋で、どのように配列を変化させて進化してきたのかを調べること。分子系統解析によって、生物や遺伝子、タンパク質が進化してきた道筋を、樹木が根から幹、そして枝へと分岐していく様子になぞらえて描くことができます。これを分子系統樹と呼び、根の位置が起源に相当し、枝の端が現在に相当します。
※6 最古の共通祖先
ほぼすべての現存地球生物が持つ核酸塩基配列やタンパク質アミノ酸配列の種間比較から作成された分子系統樹から、すべての地球生物は一つの共通祖先生物から進化してきたことがわかります。この共通祖先生物のことを「全生物の最後の共通祖先」と呼びます。全生物の最後の共通祖先は生命の起源、すなわち、地球上に最初に誕生した生物とは異なるので注意が必要です。ここでは、現存するすべての真正細菌の共通祖先や古細菌の共通祖先も含めた総称として最古の共通祖先と呼んでいます。
※7 触媒反応の活性化エネルギー
化学反応が進むためには、反応物が高エネルギー状態(遷移状態)を経て生成物に変化する必要があります。しかし、この遷移状態に到達するにはエネルギーが必要であり、これを活性化エネルギーと呼びます。活性化エネルギーが高いほど反応は起こりにくくなりますが、酵素は遷移状態を安定化させることで活性化エネルギーを低下させ、化学反応速度を大幅に速めます。
※8 元素同位体分析
過去の地球の気温を知る目的で、氷床コアや海底堆積物などに含まれる酸素の同位体比の分析がおこなわれています。水分子に含まれる酸素原子には、軽い酸素16と重い酸素18があり、気温が低いと軽い酸素16の方が蒸発しやすいですが、気温が高くなるにつれて、重い酸素18も蒸発しやすくなります。年代が分かっている氷床や堆積物に残る酸素16と酸素18の同位体の存在比を調べることで、当時の海水温や気温を推定できます。
※9 スノーボール・アース
全球凍結とも呼ばれ、過去に地球全体が氷に覆われたとする仮説です。約22億年前と6~7億年前に、大陸や海洋がほぼ完全に凍結し、極端な寒冷化が起こったと考えられています。この現象は、大気中の温室効果ガス(メタンや二酸化炭素)の減少と、氷の増加による太陽光の反射率の上昇が原因とされています。
※10 分子動力学シミュレーション
コンピュータを用いて原子や分子の動きを再現する計算手法です。原子や分子は常に運動しており、その動きをニュートンの運動方程式に基づいてシミュレーションすることで、タンパク質の構造変化や分子間の相互作用を解析できます。この手法は、実験では観測が難しい原子レベルの動的な現象を予測するために活用されています。
(7)論文情報
雑誌名:Protein Science
論文名:Insights into the low-temperature adaptation of an enzyme as studied through ancestral sequence reconstruction
執筆者名(所属機関名):崔 爽1、Subrata Dasgupta2、八木 創太1,2、木村 円香1、古川 龍太郎1、田上 俊輔2、赤沼 哲史1*責任著者
1. 早稲田大学人間科学学術院
2. 理化学研究所
掲載日時:2025年2月19日(水)
掲載URL:https://onlinelibrary.wiley.com/doi/10.1002/pro.70071
DOI:0.1002/pro.70071